



باکتری ها
باکتریها گروهی از موجودات تک یاخته ای ذره بینی هستند که پوشش بیرونی نسبتاً ضخیمی آنها را احاطه کرده است. این موجودات ساختار ساده ای دارند و به گروه پروکاریوت ها تعلق دارند.

باکتری ها متنوع ترین و مهم ترین میکروارگانیسمها هستند. تعداد کمی از آنها در انسان و حیوانات و گیاهان بیماریزا است. بطور کلی بدون فعالیت آنها، حیات بر روی زمین مختل می گردد. بطور یقین یوکاریوتها از موجودات زنده باکتری مانند بوجود آمده اند. نظر به اینکه باکتریها ساختمان ساده ای داشته و می توان به آسانی بسیاری از آنها را در شرایط آزمایشگاه کشت داد و تحت کنترل درآورد، میکروب شناسان مطالعه وسیعی درباره فرایندهای حیاتی آنها انجام داده اند
● تاریخچه
اینکه پروکاریوتها و یا یوکاریوتها کدام یک زودتر بر روی کره زمین ظاهر شده اند، کاملاً مشخص نیست. اما مطالعات تفاوتهای ژنتیکی بین یوباکتری ها، آرکی باکتری ها و یوکاریوت ها نشان می دهد که هر سه گروه از دنیای مشترکی مشتق شده اند. شکل باکتریها بر اساس شکل به ۶ گروه تقسیم می شود. پنج گروه اول را باکتریهای پست و گروه ششم را باکتریهای عالی گویند.
باکتریها دارای هسته بدون غشا و هستک هستند و اجزای آن در سیتوپلاسم پراکنده اند. کروموزمهای غیر مشابه و جداگانه در آنها وجود ندارد. در باکتریها، واکوئل دیده نمی شود. بیشتر آنها بدون کلروفیل هستند و دگرگشت (متابولیسم) خود را از راه شیمیوسنتز انجام می دهند. تولید مثل به دو صورت جنسی (آمیختگی) و غیر جسمی و جوانه زدن، قطعه قطعه شدن و تقسیم دوتایی صورت می گیرد.
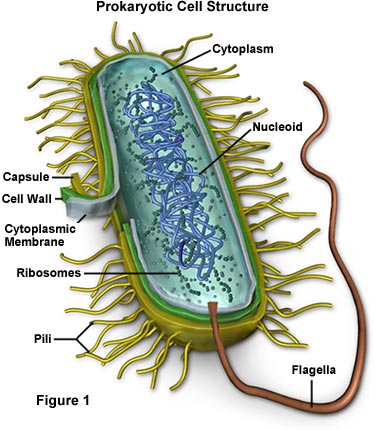
● ساختار باکتریها
پوشینه در بعضی از باکتریها، غلاف ژلاتینی چسبناکی دیواره اسکلتی را احاطه کرده است که توسط باکتریها ساخته شده و به خارج ترشح می گردد و جنس پوشینه بیشتر از پلی ساکاریدها همراه با مواد دیگر است.
تاژک از واحدهای پروتئینی به نام فلاژین تشکیل شده و قابل ترمیم بوده و وسیله حرکت باکتری هستند. معمولاً طول آن چند برابر طول باکتری است. آرایش تاژک در باکتریهای تاژکدار بصورت تک تاژکی، دو تاژکی، چند تاژکی سطحی است.
تار به دو صورت جنسی و چسبنده وجود دارد و در عمل تحرک بی تأثیر است.
دیواره در بیرون غشای پلاسمایی بوده و سبب استحکام باکتری شده و به آن شکل می دهد. وجود دیواره برای رشد و تقسیم باکتریها لازم است.
غشای سیتوپلاسمی به صورت پرده نازکی در داخل دیواره باکتری قرار دارد و متشکل از مولکولهای چربی و پروتئینی است.
مزوزومها از فرورفتگی غشای سیتوپلاسمی به درون سیتوپلاسم حاصل می شود و اغلب در محل تقسیم دیواره وجود دارند و در عمل تقسیم DNA، تقسیم یاخته ای و تبدیل باکتری به هاگ دخالت می کنند
.
اجزای سیتوپلاسم ریبوزمها مواد ذخیره ای ماده زمینه کروماتومور ماده ژنتیکی که DNA آنها غالبا به صورت یک کروموزوم تاخورده و بهم فشرده است
لویی پاستور (۲۷ دسامبر ۱۸۲۲ – ۲۸ سپتامبر ۱۸۹۵) میکروب شناس و شیمی دان فرانسوی در «دل» (Dole) منطقه ای در ناحیه «ژرا» (Jvra) در کشور فرانسه به دنیا آمد.
عمده ترین دلیل شهرت وی کشف شیوه جلوگیری از ترشیدگی شراب و شیر بود که به «پاستوریزاسیون» شهرت یافت. پدر لویی یعنی «ژان پاستور» به شغل دباغی اشتغال داشت. وی همچنین یکی از کهنه سربازان جنگ های دوران ناپلئون بود. از نظر مذهبی لویی تا پایان عمر یک کاتولیک مومن باقی ماند. (نکته جالب اینجاست که نام پاستور «Pastevr» به زبان فرانسه به معنی «کشیش پروتستان» و «چوپان» است. ) آزمایشات و تحقیقات لویی پاستور باعث تایید «تئوری نقش میکروب ها در بروز بیماری ها» شد. به علاوه وی اولین واکسن هاری را نیز کشف کرد. نام پاستور در کنار «فردیناند کوهن» و «رابرت کوخ» به عنوان بنیان گذاران باکتری شناسی ثبت شده است. همچنین پاستور اکتشافات متعددی در علم شیمی از خود به جای گذاشته، که مهمترین آنها کشف «عدم تقارن مولکولی کریستال ها» بوده است.
استعداد پاستور ابتدا توسط مدیر دانشگاه محل تحصیل اش کشف شد. این فرد پس از آن به پاستور توصیه کرد تا شانس خود را برای پذیرش در دانشسرای عالی امتحان کند، اتفاقی که با موفقیت همراه بود. پاستور پس از فارغ التحصیلی از دانشسرای عالی، مدت کوتاهی به عنوان دبیر فیزیک در دبیرستان «دیون» (۱۸۴۸) مشغول به کار شد. وی پس از آن به عنوان استاد رشته شیمی در دانشگاه استراسبورگ پذیرفته شد. جایی که استاد جوان پس از آشنایی با «ماری لوران»، دختر رییس دانشگاه در سال ۱۸۴۹ با وی ازدواج کرد. لویی و ماری صاحب پنج فرزند شدند اما تنها دو تن از آنها به سنین بزرگسالی رسیدند. خدمات علمی پاستور اثبات تئوری نقش میکروب ها در بروز بیماری ها که پیش از آن به فکر افرادی چون «گیرولامو فرانکاستورو»، «آگوستینو باسی» و «فردریش هلن» خطور کرده بود، با آزمایش های پاستور تحقق یافت. پاستور موفق شد این تئوری را تبدیل به نظریه ای قابل پذیرش برای اکثر دانشمندان عصر خود کند.
این باکتری (H. pylori) شایع ترین موجود ذره بینی است که جوامع انسانی را در بعد جهانی مبتلا به عفونت ساخته و گفته میشود بیش از ۵۰ درصد از مردم دنیا میزبان این بیماریزایی میباشند.معده انسان تنها محل اقامت مناسب این میکروب است و حتی از دوران نوزادی بعد از شیر خوارگی تا سنین کهولت، انسان میتواند ناقل این باکتری باشد.
این میکروب در محل اقامت طویل مدت خود در معده میتواند باعث بیماریهایی به صورت التهاب حاد و مزمن معده ، زخم تغذیه ای معده و اثنی عشر با عواقب خطر ناکشان از جمله خونریزی حاد و مزمن جهاز هاضمه و سرطان معده شود.در اکثر مردم این عفونت به صورت بدون علائم باقی می ماند و لذا ناقلین آن سبب سرایت آن به افراد دیگر چه در هسته فامیلی و چه در اجتماعات متراکم انسانی مانند خوابگاه میشوند.
علت عدم بروز علائم بالینی در تمام اشخاص آلوده مشخص نیست ونوع سویه باکتری ، تغذیه و سایر عوامل اجتماعی مسئول آن شناخته شده اند.
رابطه علی آن با سرطان معده در اشخاص مبتلا به عفونت به اثبات رسیده است لذا در جوامع در حال توسعه این عفونت شایع ترین علت مرگ در نتیجه سرطان معده محسوب میگردد. نشان داده شده است که این عفونت در قالب حیوانی با تلقیح واکسن قابل درمان است ولی واکسن نوع انسانی هنوز در دسترس نیست.بر اساس مطالعات میتوان گفت شیوع این عفونت در ایران بسیار بالا است.
● محیط زیست :H .pylori
این باکتری در محیط طبیعی به شکل نیم دایره یا به شکل S و در محیط آزمایشگاهی به شکل میله ای مشاهده شده است و به همین دلیل این نام به آن اطلاق گردیده است .محیط زیست این باکتری در سطح مخاطی اپیتلیوم معده به خصوص در قسمت پیلور میباشد.
به اثبات رسیده است که این میکروب در نسوج پوششی معده در اثنی عشر و به ندرت در مری و مقعد ودر مخاط معده مشاهده شده به نظر میرسد که این میکروب در دوران طفولیت وارد بدن شده و برای زندگی طویل مدت در نقاطی که فاقد میکروب های طبیعی هستند خود را تطبیق میدهد.
هلیکو باکتر پیلوری گاهی از بزاق دهان و جرم دندانها جدا شده است. مدفوع یک منبع احتمالی برای آن محسوب میشود،البته محققین به این نتیجه رسیده اند که دفع باکتری در مدفوع زیاد نیست.
انسان میزبان طبیعی این باکتری است و بر اساس یک سری مطالعات پیشنهاد شده که حیوانات میتوانند به عنوان مخازنی برای باکتری عمل کنند . این باکتری قابل انتقال از حیوان به انسان است. در میمونها به خصوص در میمون هندی به نام ریسوس ،خوک،عنتر و گربه های خانگی و بعضی حیوانات اهلی گزارش شده است.
انتقال این باکتری از شخص به شخص در صورت رعایت نکردن بهداشت فردی امکان پذیر است.
● درمان:
این باکتری یک ارگانیزم بسیار مقاوم به درمان است و موفقیت در درمان نیاز به تجویز همزمان دو یا چند دارو دارد.آموکسی سیلین بطور گسترده در جهت در مان به خصوص در ترکیب با بیسموت استفاده شده است.
درمان سه دارویی هم فوائد داخل معده ای و هم فعالیت سیستمیک داشته است .داروهای داخل معده ای موثر شامل بیسموت، تتراسایکلین، آموکسی سیلین، کلاریترومایسین و فورازولیدون هستند.
در صورت ابتلا به این باکتری دوره درمانی ودارویی خود را با جدیت دنبال نمایید زیرا در صورت سهل انگاری عود مجدد بیماری گزارش شده است .
● رنگ آمیزی گرم
رنگ آمیزی گرم یکی از مهمترین ومتداولترین روش های رنگ آمیزی در باکتری شناسی است که اولین بار توسط کریستین گرم ابداع شد. دراین رنگ آمیزی باکتری ها بر مبنای رنگ باکتری پس ازرنگ آمیزی به دودسته گرم مثبت و گرم منفی تقسیم می شوند. رنگ باکتری پس ازرنگ آمیزی به توانایی حفظ رنگ اول وبه عبارتی به ساختمان دیواره سلولی باکتری بستگی دارد. دررنگ آمیزی گرم باکتری های گرم مثبت پس ازرنگ آمیزی به رنگ بنفش وباکتری های گرم منفی به رنگ قرمز مشاهده می شود.
گرچه هر دو گروه یعنی باکتری های گرم مثبت و منفی دارای دیواره می باشند ولی فرق بین این دو گروه مربوط به خواصی است که در ساختمان دیواره سلولی آنها وجود دارد. اساس ساختمان در دیواره سلولی باکتری های گرم مثبت یک لایه ضخیمی است از پپتیدوگلیکان*[۱]، ولی در باکتری های گرم منفی ضخامت آن به حداقل می رسد.
در عمل باکتریهایی که دارای خواص یکسانی باشند بندرت یافت می شوند، حتی باکتریهایی که از یک سلول منشا می گیرند ممکن است از نظر یک یا چند صفت با یکدیگر متفاوت باشند. این تفاوتها نتیجه تغییراتی است که به علت جهش ژنی یا موتاسیون در سلولهای باکتریایی پدید می آید. این باکتریهای تغییر یافته ، موتانت Mutant نامیده می شوند که از نظر بعضی از خواص نظیر ساختمان آنتی ژن ، حساسیت در مقابل آنتی بیوتیکها و ... با سایر باکتریهای مشابه اختلاف دارند.
سهولت تغییرپذیری در باکتریها مربوط به سرعت تقسیم آنهاست. زمان تقسیم یا مدت زمانی که برای تولید یک سلول جدید در باکتریها لازم است، حدود ۲ دقیقه و در مورد انسان ۲۰ سال است. مثلا یک سلول باکتری در مدت ۱۸ ساعت ۵۴ نسل بوجود می آورد. درحالیکه برای ایجاد همین تعداد نسل انسان بیش از ۱۰۰۰ سال زمان لازم است. پس جهش ژنی در باکتریها نسبت به موجودات عالی خیلی سریع و قابل ملاحظه است.
تفاوت یوکاریوتها با باکتریها
در کره خاکی تنها دو نوع سلول توسط کلیه ارگانیسمهای زنده تولید می شود. سلولهای پروکاریوت (یا هسته ابتدایی). در این گروه هسته ، فاقد غشا است و شامل کلیه باکتریهاست. پروکاریوتها شامل یو باکتریها (باکتریهای حقیقی) و آرکئی باکترها (باکتریهای قدیمی) است. اما گروه دیگر یوکاریوتها هستند که دارای غشای هسته و هسته حقیقی می باشند. اینگونه هسته در تمام ارگانیسمهای دیگر مانند Algae (جلبکها) Fungi (قارچها) ، پروتوزوئرها (protozoa) و گیاهان (Plant) و جانوران (Animals) یافت می شود. پاتوژنهای انسانی تنها در میان یوباکتریها یافت می شوند
دوباره می گوییم که کروموزوم باکتریایی یک مولکول دی ان ای دی ان ای دو رشته ای است که در یک لوپ بزرگ محاط می شود چون تنها یک کپی از این مولکول در هر سلول وجود دارد باکتری در شرایط هاپلویید به بقای خود ادامه می دهد.باکتری ها غشای هسته ای که دی ان ای آن ها را احاطه کنند ندارند زیرا فاقد هیستون اند و ماده ای شبیه هیستون دارند که به پیشرفتگی هیستون نیست.باکتری اگر مثل یک سلول یوکاریوت هسته ای با غشا می داشت آن هنگام ارتباط بین تنها دی ان ای باکتری با خارج از هسته سخت یا منحل میشد و باکتری نمی توانست اصلا به حیات ادامه دهد یا به طور کامل تشکیل شود ولی در سلول های یوکاریوتی به دلیل تراکم بالای دی ان ای و وجود هیستون ها غشای دیواره هسته مشکلی ایجاد نکرده و ارتباطی غیر مستقیم را با سایر نقاط سلول خاص یوکاریوتی برقرار می کند اما دی ان ای در باکتری ها و پروکاریوت ها در ارتباط مستقیم با سایر اجزا قرار دارد و این فرآیند نیز سرعت تکثیر باکتری را افزایش می دهد
● مشخصات سلول باکتری
اکثر باکتریها پوشش سلولی (cell envelope) تولید می کنند که شامل غشای پلاسمایی ، دیواره سلولی (cell wall) و پروتئینها و پلی ساکاریدهای تشکیل دهنده آن می باشد. بعضی از باکتریها کپسول یا لایه چسبنده تولید می کنند. فیلامانهای خارجی (فلاژل و پیلی) ممکن است در باکتریها بوجود آید. دیواره سلولی ، ساختمان سخت و مقاومی است که پروتوپلاست را احاطه کرده و آن را از آسیب فیزیکی و شرایط کاهش فشار اسمزی محیط خارج حفاظت می کند. معمولا به باکتری اجازه می دهد تا در برابر سطح وسیعی از شرایط محیطی ایستادگی کند پروتوپلاست از غشای سیتوپلاسمی و محتویات آن تشکیل شده است.
از نظر محتویات سلولی ، باکتریها سلولهای ساده ای هستند. ساختمان اصلی سیتوپلاسم آنها شامل شبکه فیبریلی کروماتین مرکزی یا نوکلئوتید (Nucleoid) می باشد که توسط سیتوپلاسم بی شکل حاوی ریبوزوم ها احاطه شده است. اجسام انکلوزیون سیتوپلاسمی یا گرانولهای ذخیره انرژی ، بسته به گونه های باکتری ماهیت شیمیایی متفاوتی دارند و مقدار آنها به مرحله رشد و محیط بستگی دارد. بعضی از ساختمانهای سلولی از قبیل آندوسپورها فقط به تعداد کمی از باکتریها محدود می شوند.
● طبقه بندی باکتریها
▪ باکتری های پست
این باکتریها تک یاخته ای بوده و اگر کروی یا بیضوی باشند، کوکوس و اگر میله ای شکل یا دراز باشند، باسیل و اگر خمیده باشند ویبریون و چنانچه مارپیچی شکل و غیرقابل انعطاف باشند، اسپریل و اگر فنری و قابل انعطاف باشند، اسپیروکت نامیده می شوند.
● باکتریهای عالی یا رشته ای
این باکتریها رشته مانند و اغلب غلاف دار هستند و اغلب اوقات شاخه های حقیقی ایجاد کرده ، میسلیوم تشکیل می دهند و چون تشکیلات منشعب ایجاد می کنند، لذا اکتینومیست نامیده می شوند. بنابراین باکتریها از نظر شکل به ۶ گروه گرد ، دراز ، خمیده ، مارپیچی ، فنری و منشعب تقسیم می شوند.
● اجزای ساختمانی باکتریها
▪ فلاژلها (Flagella)
فلاژلها ، فیلامانهای پروتئینی به طول و قطر یکنواخت می باشند و موجب تحرک شبیه به شنای سریع و مستقل اغلب باکتریها پاتوژنیک می گردند فلاژل در سه قسمت فیلامان ، قلاب و جسم پایه تشکیل شده است. پایه فلاژل در غشای پلاسمایی قرار گرفته است. لنگرگاه و تعداد فلاژل در باکتریها فرق خواهد کرد.
● فیمبریاها
فیمبریاها که پیلی هم نامیده می شوند، فیبریلهای شبیه مو هستند به اندازه ۰.۰۰۴ تا ۰.۰۰۸میکرون هستند. این ارگانل با میکروسکوپ الکترونی در سطح باکتریهای مختلف قابل رویت هستند. آنها مستقیم تر ، نازکتر و کوتاهتر از فلاژلها هستند. این رشته ها در غشای پلاسمایی سلول میکروبی لنگر می اندازد.
● هسته باکتری
هسته سلول را میتوان بعد از رنگ آمیزی اختصاصی با میکروسکوپ نوری مشاهده کرد. در مقایسه با سلولهای عالی مواد ژنتیکی باکتریها و سایر سلولهای پست پراکنده ، ساده و بدون پوشش و کروموزوم حلقوی است غشای هسته وجود ندارد و کروموزوم به مزوزوم فرورفته در غشای سیتوپلاسمی چسبیده است. در سالهای اخیر پروتئینهای شبیه هیستون در باکتریها کشف شده است که احتمالا نقش مشابه هیستونها را در کروماتینهای سلولهای یوکاریوت ایفا می کنند.
● سیتوپلاسم
بیش از ۵۰ درصد پروتئین سلول در سیتوپلاسم قرار دارد و آنزیمهای متابولیسمی راههای گلیکولیز و بسیاری از آنزیمهای چرخه کربس ، انواع کاتالازها ، دهیدروژنازها ، و مواد حد واسط چرخه های متابولیکی در سیتوپلاسم وجود دارد. روابط اتمی ، یونی و الکترونی بین ترکیبهای مختلف سیتوپلاسمی با نظم خاص فعالیتهای حیاتی را ظاهر می سازد.
●پوشش سلول (Cellenvelope)
▪ کپسول و لعاب (Capsoles)
قدرت بیماری زایی پاتوژنها اغلب با تولید کپسول همراه است. باکتریهای کپسول دار در محیط جامد ، کلنیهای مخاطی (Mucoid) یا صاف (Smooth) می سازند. در مقابل باکتریهای فاقد کپسول کلنیهای خشن (Rough) دارند. اگر باکتری قدرت کپسول سازی خودش را از دست بدهد در مقابل قدرت ویرولانس (بیماریزایی) خود را از دست داده و در مقابل دستگاه ایمنی بدن میزبان تاب مقاومت نخواهد داشت.
● دیواره سلولی
دیواره سلولی باکتریها بی نهایت پیچیده است و لایه سفت و سختی را در اطراف باکتریها ایجاد می کند که سلول را از گسیختگی و متلاشی شدن در مقابل فشار اسمزی خارج سلول محافظت می کند. همچنین دیواره محل تجمع عوامل آنتی ژن می باشد که باکتریها را توسط این آنتی ژنها از هم تمیز می دهند. باکتریها با روش رنگ آمیزی گرم (Gram stain) به دو دسته تقسیم می شوند.
گرچه هر دو گروه یعنی باکتریهای گرم مثبت و منفی دارای دیواره می باشند ولی فرق بین این دو گروه مربوط به خواصی است که در ساختمان دیواره سلولی آنها وجود دارد. اساس ساختمان در دیواره سلولی باکتریهای گرم مثبت یک لایه ضخیمی است از پپتیدوگلیکان (Poptidoglycan) ، ولی در باکتریهای گرم منفی ضخامت آن به حداقل می رسد.
● غشای سیتوپلاسمی
غشای سیتوپلاسمی غشای داخلی نیز نامیده می شود. غشای سیتوپلاسمی باکتریها مشخص بوده و از فسفو لیپید و پروتئین ساخته شده است. این غشا در پروکاریوتها از غشای سیتوپلاسمی در یوکاریوتها به علت نداشتن استرول متمایز می شود. چین خوردگیهای غشای سیتوپلاسمی به درون سلول ساختارهای ویژه ای به نام مزوزوم ایجاد می کند که کروموزومهای باکتریها به مزوزومها متصل هستند. غشا همچنین به عنوان یک سد اسمزی برای سلول عمل می کند و دارای سیتوپلاسم انتقال دهنده برای مواد محلول است و انتقال تولیدات سلولی را در مقابل با محیط خارج سلولی تنظیم می کنند.
● تولیدمثل باکتری
باکتریها به روشهای تقسیم مستقیم ، آمیختگی ، قطعه قطعه شدن یا بوسیله کنیدی و همچنین جوانه زدن تکثیر می یابند. برخی باکتریها توانایی ایجاد هاگ درونی را دارند. هاگ سبب مقاومت باکتری در برابر عوامل نامساعد محیط می شود. هر باکتری فقط یک هاگ می سازد و از هر هاگ یک باکتری بوجود می آید.
به تازگی جمعی از مایکروبایولوژیست ها در یکی از دانشگاههای انگلیس یکی از باکتری های موجود در شکلات را که از شکر تغذیه می کند به وسیله اضافات شکلاتهای یک کارخانه پرورش دادند و سپس یک باطری هیدروژنی را با هیدروژن تولید شده از فعالیت باکتری ها شارژ کرده و ثابت کردند که می توان از شکلات انرژی الکتریکی دریافت کرد.
این تیم تحقیقاتی باکتری اسچریچیا کولی (Escherichia coli) موجود در شکلات را با کارامل رقیق شده تغذیه کردند، باکتری مورد نظر شکر را مصرف و هیدروژن تولید کرد.
سپس محققین هیدروژن حاصل شده از باکتری را برای شارژ یک باطری هیدروژنی استفاده کردند و باطری هیدروژنی نیز پس از شارژ الکتریسیته کافی برای راه انداختن یک دستگاه خنک کننده کوچک را تولید کرد.
این کشف تازه راه مصرف فوق العاده ای را برای استفاده از اضافات و زایده های کارخانه های شکلات سازی ایجاد خواهد کرد.
جالب اینجاست که کار این باکتری تنها با یک بار مصرف شکر و تولید هیدروژن تمام نمی شود و می تواند همچنان درعرصه تامین سوخت هیدروژنی فعال باشد. دانشمندان باکتری را در ظرف مخصوصی حاوی هیدروژن و مایع زاید حاصل شده از روند تبدیل شکر به هیدروژن قرار می دهند و دوباره آنزیم تولید کننده هیدروژن را در آنها فعال میکنند.
محققین برای استفاده مجدد از باکتری ها ، گاز هیدروژن را به الکترون های تشکیل دهنده آن تجزیه می کنند سپس الکترون های تولید شده را با الکترون های فلز پلادیوم در محلول هیدروژن و مایع زاید حاصله از فعالیت باکتری ها قرار می دهند تا الکترون ها با هم واکنش شیمیایی انجام دهند.
این واکنش موجب می شود تا پلادیوم از محلول جدا شده و به باکتری بچسبد و در پی این عمل باکتری برای استفاده مجدد حفظ می شود.
ژنتیک باکتریایی
کروموزوم باکتریایی یک ملکول DNA دو رشته ای است که در یک لوپ بزرگ محاط می شود. چون تنها یک کپی از این ملکول در هر سلول وجود دارد، باکتری در شرایط هاپلویید به حیات ادامه می دهد. باکتری ها غشای هسته ای که DNA آنها را احاطه کند، ندارند.
این فصل تلاش ندارد به تمامی جزئیات ژنتیک باکتریایی مثل همانندسازی(replication)، رونویسی (transcription) و ترجمه (translation) بپردازد. به این عناوین به طور گسترده در آموزش ژنتیک پرداخته می شود. درعوض این فصل به مکانیسم های تبادل اطلاعات ژنتیک باکتریایی می پردازد.همانطور که می دانید پروکاریوت ها این مکانیسم ها را به صورت ابتدایی دارند زیرا آنها درگیر جوش خوردن جنسی (sexual union) با سایر باکتری ها نمی شوند. باکتری ها همانندسازی ژنی انجام می دهند، یک نسخه عینا مشابه از ژنومشان را ایجاد می کنند وسپس به دو بخش تقسیم می شوند در حالی که هر قسمت یک نسخه دارد (تقسیم دوتایی/binary fission). سلول های ارگانیسم های تکامل یافته تر (یوکاریوت ها) مجموعه ای از گامت ها از هر والد ایجاد می کنند بنابراین تنوع ژنتیکی را تضمین می کنند. پس چگونه موجودات بدون جنسیت (sexless) دستخوش تغییر ژنتیکی می شوند که برای بقا بسیار ضروری است ؟
یک مکانیسم، موتاسیون ساده است. هرچند نادر است که یک موتاسیون نقطه ای ساده، ارگانیسم را به شیوه ای مفید تغییر دهد. جهش های نقطه ای معمولا منجر به بی معنی بودن یا معنی اشتباه می شوند (ایا این معنی دارد؟). چهار روش وجود دارد که در آن باکتری می تواند قطعات ژنتیکی را مبادله کند:1- ترانسفورماسیون(transformation)، 2- ترانس داکشن (transduction)، 3- کنژوگیشن (conjugation) (so much for celibacy)، 4- ترانس پوزون (transposon)
بقا = تغییرات
تبادل مواد ژنتیکی به اشتراک گذاشتن ژن هایی را ممکن می کند که پروتئین هایی مانند آن هایی که مقاومت آنتی بیوتیکی ایجاد می کنند، اگزوتوکسین ها،آنزیم ها،و سایر ویرولانس فاکتورها (پیلی،تاژک و کپسول) را کد می کنند. دانشمندان می توانند از این مکانیسم های تبادل برای مهندسی ژنتیک و تعیین نقشه کرومزومی استفاده کنند. به خواندن ادامه دهید ... ولی تنها اگر بیش از 21 سال سن دارید.
ترانسفورماسیون
قطعات برهنه DNA یک باکتری که در هنگام لیز سلول رها شدند به دیواره سلولی یک باکتری دیگر می چسبند. باکتری گیرنده باید باکفایت (competent) باشد به این معنی که بر روی دیواره سلولی خود ساختارهایی داشته باشد که بتواند به DNA متصل شود و آن را به داخل سلول جذب کند. باکتری های گیرنده باکفایت (competent) معمولا از همان گونه های دهنده هستند. اگر همولوژی (homology) کافی بین رشته ها وجود داشته باشد DNA وارد شده به ژنوم گیرنده ملحق می شود (دلیلی دیگر برای اینکه چرا این انتقال تنها می تواند بین خویشاوندان نزدیک باکتریایی صورت گیرد).
مثال معروف از این گونه تبادلات،آزمایشی است که در سال 1928 توسط فردریک گریفیت (Frederick Griffith) اجرا شده است.او از باکتری استوپتوکوکوس پنومونیه (Streptococcus pneumoniae) استفاده کرد که بر اساس تفاوت هایی در کپسول سلولی به سرده های متفاوت بسیاری طبقه بندی شده است. گریفیت از پنوموکوک صاف و کپسول دار که باعث عفونت شدید و مرگ در موش می شود و پنوموکوک خشن بدون کپسول که موش را نمی کشد استفاده کرد. .........؟........ (آن ها بی دردسرند و نمی توانند کک را بکشند). گریفیت باکتری های مضر صاف کپسول دار را با گرما کشت و آن ها را به همراه باکتری های بی دردسر بدون کپسول خشن زنده به موش تزریق کرد. بنگر و ببین موش مرد، و وقتی او باکتری ها را از خون کشت داد تنها توانست پنوموکوک صاف کپسول دار زنده پیدا کند. ژن کد کننده کپسول از باکتری های کشته شده با گرما آزاد شده و به باکتری های بدون کپسول خشن زنده ملحق شده بود. بنابراین باکتری های خشن به باکتری های صاف کپسول دار کشنده تبدیل شده بودند.
امروزه دانشمندان به طور گسترده از این روش برای وارد کردن DNA نو ترکیب و برای نقشه برداری ژن روی کروموزوم استفاده می کنند. این روش می تواند در نقشه برداری استفاده شود چون فراوانی جابجایی هایی (transformation) که منجر به انتقال دو صفت می شود با فاصله آن ها در ژنوم مرتبط است. هرچقدر آن ها به هم نزدیک تر باشند با احتمال بیشتری با هم مبادله خواهند شد.
ترانسداکشن
ترانس داکشن زمانی اتفاق می افتد که یک ویروس که باکتری را آلوده می کند (که باکتریوفاژ نامیده می شود)، بخشی از DNA باکتریایی را از یک باکتری به دیگری حمل کند. برای فهم این موضوع بگذارید لحظاتی از موضوع منحرف شویم و درباره باکتریوفاژ صحبت کنیم.
شکل 1-3. باکتریوفاژها (bacteriophages) در داشتن یک پوشش پروتئینی به نام کپسید که یک ملکول DNA یاRNA را احاطه می کند همانند اکثر ویروس ها هستند. باکتریوفاژها تقریبا مشابه عنکبوت هستند با گردن های دراز و باریک.
فاژ به وسیله رشته های دمی اش به گیرنده ای اختصاصی بر روی سطح سلول باکتریایی متصل می شود،این عمل رونشینی (Adsorption) نام دارد. سپس فاژ متحمل نفوذ می شود. با شباهت بسیاری به یک عنکبوت که چمباتمه می زند و نیشش را فرو می کند، فاژ لوله توخالی زیر غلاف گردنش را از میان دیواره سلول و غشای سیتوپلاسمی رد می کند. DNA داخل سر از میان لوله به داخل باکتری تزریق می شود.
شکل 2-3. به دنبال رونشینی و نفوذ، DNAی تزریق شده برای رونویسی DNA فاژ به RNA پیامبر(mRNA) ، RNA پلیمراز میزبان باکتریایی را تصاحب می کند. کپسیدهای جدید، DNA و آنزیم ها تشکیل شده اند و سلول باکتریایی با فاژهای جدید پر می شود. در جایی سلول نمی تواند ذرات بیشتری را نگه دارد و لیز می شود و فاژها آزاد می گردند.
برای پیچیده تر کردن مطلب، دو نوع فاژ وجود دارد، فاژهای کشنده (virulent phages) و فاژهای معتدل (temperate phages). فاژهای کشنده همانند آنچه در شکل 2-3 نشان داده شده رفتار می کنند. باکتری را آلوده می کنند، تکثیر می شوند، سپس باکتری را لیز می کنند و می کشند. ازطرف دیگر فاژهای معتدل خلق و خوی خوبی دارند و باکتریی که آلوده کردند را بلافاصله لیز نمی کنند. فاژ معتدل همانند فاژ کشنده رونشینی و نفوذ می کند اما بعد، به جای رونویسی، DNA آن به کروموزوم باکتریایی ملحق می شود. سپس DNA منتظر یک دستور برای فعالسازی می ماند.
شکل 3-3. ژنوم ترکیب شده ی فاژ معتدل، پروفاژ(prophage) نامیده می شود. باکتری هایی که یک پروفاژ ترکیب شده با کروموزومشان دارند لیزوژنتیک (lysogenic) نام دارند زیرا گاهی پروفاژ سرکوب شده می تواند فعال شود. هرگاه فعال شود، پروفاژ تولید فاژهای جدید را آغاز می کند، و چرخه ای را شروع می کند که با لیزسلول باکتریایی خاتمه می یابد. بنابراین فاژهای معتدل با وجود خلق و خوی خوب مانند بمب های ساعتی کوچک ژنتیکی هستند.
ایمنی لیزوژنتیک (lysogenic immunity) اصطلاح استفاده شده برای توصیف توانایی یک باکتریوفاژ ترکیب شده (پروفاژ) برای جلوگیری از آلودگی بعدی توسط فاژ مشابه می باشد. اولین فاژ معتدل آلوده کننده یک باکتری، پروتئین سرکوبگر تولید می کند. این سازش "بقای اصلح" تضمین می کند که اولین فاژ معتدل تنها اشغال کننده باکتری باشد.
اکنون که با باکتریوفاژها آشنا شدیم، بگذارید بحث کنیم چگونه این فاژها می توانند DNA باکتریایی را از یک باکتری به دیگری منتقل کنند. این فرآیند ترانس داکشن (transduction) نامیده می شود. همانطورکه دو نوع فاژ وجود دارد، دو نوع ترانس داکشن نیز موجود است. فاژهای کشنده در ترانس داکشن عمومی (generalized transduction) و فاژهای معتدل در ترانس داکشن اختصاصی (specialized transduction)قرار می گیرند.
ترانس داکشن عمومی
ترانس داکشن عمومی به صورت زیر رخ می دهد. پس از نفوذ فاژ به باکتری میزبان، DNA فاژ به کپسید و آنزیم ها رونویسی، همانند سازی و ترجمه می شود. در همین زمان DNA باکتریایی سرکوب و سرانجام نابود می شود. گاهی قطعات DNA باکتریایی سالم می مانند. اگر این قطعات هم اندازه DNA فاژ باشند می توانند به طور تصادفی داخل سر کپسید فاژ قرار گیرند. به دنبال لیز سلول و رها شدن فاژها، فاژی که در سرش DNA باکتریایی دارد، می تواند باکتری دیگری را آلوده کند. فاژ قطعه ی DNA باکتریایی که بطور تصادفی حمل می کند را تزریق خواهد کرد. اگر بین رشته تازه تزریق شده و ژنوم باکتریایی گیرنده قدری همتایی (همولوژی) وجود داشته باشد امکان دارد قطعه به آن ملحق شود. ژن واقع بر روی آن قطعه می تواند پروتئینی را کد کند که گیرنده آن را در ابتدا نداشته است، مانند پروتئینی که یک آنتی بیوتیک را غیر فعال می کند. در ترانس داکشن عمومی، باکتریوفاژ تنها DNA باکتریایی را حمل می کند، بنابراین سلول گیرنده زنده خواهد ماند (زیرا هیچ ژن ویروسی که همانند سازی و لیز را کد کند موجود نیست). این نوع انتقال ژنتیکی از ترانسفورماسیون مؤثرتر است زیرا قطعه DNA منتقل شده، توسط کپسید فاژی که آن را نگه می دارد از نابودی درهنگام جابجایی حفظ می شود.
شکل 4-3. ترانس داکشن عمومی
- رونشینی و نفوذ رخ می دهد. DNA ویروسی به شکل خط باریکی کشیده شده، و DNA حلقوی باکتریایی به شکل دایره ای ضخیم رسم شده است.
- تخریب DNA باکتریایی تعدادی قطعه سالم (ضخیم) برجای می گذارد. DNA فاژ همانندسازی کرده است.
- کپسیدها ترجمه و بسته بندی شده اند. کپسید میانی حاوی قطعه ای از DNA باکتریایی است.
- لیز سلولی رخ می دهد، فاژها از جمله فاژ دارای DNA باکتریایی آزاد می شوند.
ترانس داکشن اختصاصی
ترانس داکشن اختصاصی توسط فاژهای معتدل رخ می دهد. به یاد آورید که فاژ معتدل نفوذ می کند و سپس DNA آن به کروموزوم باکتریایی ملحق می شود. فاژ از این پس پروفاژ نامیده می شود و باکتری هم اکنون لیزوژنیک است (شکل 3-3). به طور معمول پروفاژ فقط بدون انجام کاری منتظر می ماند، اما سرانجام می تواند فعال شود. اگر فعال شد، DNA پروفاژ ازکروموزوم باکتریایی بریده شده و سپس به یک کپسید همانند سازی، ترجمه و بسته بندی می گردد. گاهی در برش خطا وجود دارد و قطعه ای از DNA باکتریایی که در یک طرف از پروفاژ قرار دارد با DNA فاژ بریده، همانندسازی و بسته بندی خواهد شد. این عمل می تواند منجر به انتقال آن قطعه از DNA باکتریایی به باکتری های دیگر شود.
شکل 5-3. ترانس داکشن اختصاصی توسط فاژ لامبدا (lambda) در اشرشیا کلای (ٍٍٍEscherichia coli) اتفاق می افتد. محل درج پروفاژ لامبدا در میان ژن اشرشیا کلای برای سنتز بیوتین و سنتز گالاکتوز قرار دارد. اگر خطای برش رخ دهد، ژن بیوتین (BIO) یا ژن گالاکتوز (GAL) (ولی نه هر دو، زیرا قطعه DNA بریده شده دارای طول معینی می باشد) توسط DNA فاژ حمل و بسته بندی می شود. بنابراین ژن سنتز بیوتین اکنون می تواند به یک باکتری دیگر که این قابلیت را ندارد منتقل شود. شما مکررا راجع به این فرم انتقال ژن خواهید شنید؛ این روش کنورسیون لیزوژنیک (lysogenic conversion) نامیده می شود. برای مثال ژن اگزو توکسین کورینه باکتریوم دیفتریا (Corynebacterium diphtheria) توسط کنورسیون لیزوژنیک کسب شده است.
کانژوگیشن
کنژوگیشن سکس باکتریایی در بهترین حالت است: پرشور و جدی! در کنژوگیشن DNA مستقیما توسط تماس سلول با سلول منتقل می شود، و منجر به تغییر بسیار کارآمد در اطلاعات ژنتیکی می شود. این تبادل می تواند بین باکتری های غیر خویشاوند (unrelated) رخ دهد و مکانیسم عمده برای انتقال مقاومت به آنتی بیوتیک می باشد. برای اینکه کنژوگیشن اتفاق بیفتد، یک باکتری باید پلاسمید خود منتقل شونده (self-transmissible plasmid) داشته باشد، که پلاسمید F هم نامیده می شود (به خاطر fertility، نه کلمه دیگری!). پلاسمیدها ملکول های DNA دو رشته ای حلقوی هستند که بیرون کروموزوم قرار دارند و می توانند ژن های ضروری برای انجام فرآیند کنژوگیشن را کد کنند. باکتری هایی که F پلاسمید را حمل می کنند سلول های F(+) نامیده می شوند. در کنژوگیشن یک سلول F(+) دهنده پلاسمید F خود را به یک سلول f(-) گیرنده منتقل می کند به این ترتیب گیرنده را F(+) می کند.
شکل 6-3. پلاسمید خود منتقل شونده (پلاسمید F) ژنی دارد که آنزیم ها و پروتئین هایی را کد می کنند که آلت تناسلی و به عبارت دیگر پیلی جنسی را شکل می دهند.
این ساختار پروتئینی بلند از سطح سلول باکتری F(+) دهنده بیرون می زند و به غشای سلولی باکتری گیرنده متصل می شود و نفوذ می کند (این در نهایت جالب می شود!). اکنون که یک پل کنژوگه کننده (conjugal bridge) ایجاد شده، یک نوکلئاز یک رشته از DNA پلاسمید F را قطع می کند، و این تک رشته ی DNA از طریق پیلی جنسی (پل کنژوگه کننده) به باکتری گیرنده منتقل می شود.
شکل 7-3. وقتی یک رشته DNA از طریق پل کنژوگه کننده منتقل شد، رشته باقیمانده با بازهای نوکئوتیدی جدید جفت می شود (خط نقطه چین). اتفاق مشابهی برای رشته ای که به سلول دیگر منتقل می شود می افتد. در پایان جوش خوردن جنسی پل کنژوگه کننده تخریب می شود و هر دو باکتری پلاسمید F دو رشته ای حلقوی خواهند داشت. سلول گیرنده F(-) اکنون F(+) است.
شکل 8-3. به ندرت پلاسمید F خارج کروموزومی با کروموزوم باکتریایی مجاور ترکیب می شود. در روشی بسیار مشابه با آنچه باکتریوفاژ معتدل انجام می دهد. سلول باکتریایی از این پس سلول Hfr نامیده می شود (فراوانی بالای کروموزوم های نوترکیب). این ترکیب شدن می تواند به دو مکانیسم منحصربه فرد انتقال DNA منجر شود:
1)پلاسمید F که اکنون با کل DNA حلقوی باکتریایی همراه است با یک سلول F(-) کنژوگیشن نرمال انجام می دهد. کل کروموزوم باکتریایی شامل پلاسمید F ترکیب شده از سلول Hfr به سلول گیرنده منتقل می شود.
2)پلاسمید F ترکیب شده در سلول Hfr ممکن است از جای متفاوتی از محل ترکیب شدن بریده شود. این عمل می تواند منجر به پلاسمید F شود که اکنون قطعه ای از DNA کروموزومی را نیز دارد. این پلاسمید ها، پلاسمیدهای F' (F prime) نامیده می شوند. این کنژوگیشن 'F متناظر ترانس داکشن اختصاصی است زیرا در هر دو حالت قطعه مجاور از DNA کروموزومی به طور تصادفی برداشته می شود و می تواند به یک سلول باکتریایی دیگر منتقل شود.
برخی پلاسمیدها، پلاسمیدهای غیر خود منتقل شونده هستند. این پلاسمیدها ژن های لازم برای هدایت کنژوگیشن را ندارند. به هرحال آن ها به همراه باکتری میزبانشان همانند سازی می شوند و به منتقل شدن ادامه می دهند در حین اینکه باکتری در تقسیم دوتایی تقسیم می شود.
پلاسمیدها از نظر پزشکی فوق العاده مهم هستند.پلاسمیدهای خاص آنزیم هایی را کد می کنند که آنتی بیوتیک ها را تجزیه می کنند (پنی سیلیناز) ، یا ویرولانس فاکتورها را تولید می کنند (مثل پیلی واگزوتوکسین).
ترانس پوزون ها
شکل 9-3. ترانس پوزون ها عناصر ژنتیکی متحرک هستند. شما می توانید آنها را قطعات DNA پادار مجسم کنید. این قطعات DNA می توانند خودشان را بدون داشتن همولوژی DNA در کروموزوم دهنده وارد کنند. ترانس پوزون ها می توانند ژن های مقاومت به آنتی بیوتیک و ویرولانس فاکتورها را حمل کنند.
ترانس پوزون ها وارد DNA فاژها،پلاسمیدها و کروموزوم های باکتریایی می شوند. آن ها به طور مستقل همانند سازی نمی کنند بلکه در طی رونویسی DNA میزبانشان کپی می شوند. وقتی ترانس پوزون ها DNAیی که با آن ترکیب شده اند را ترک می کنند غالبا برش اشتباهی وجود دارد و ترانس پوزون می تواند DNA جدید را به محلی دیگر منتقل کند. اهمیت ترانس پوزون ها...در این است که یک ژن ترانس پوزون که با مقاومت دارویی خاصی مداخله می کند می تواند به پلاسمید های سرده های متفاوت باکتریایی متصل شود و منجر به پراکندگی سریع گونه های مقاوم شود.

اگر قبلا در بیان ثبت نام کرده اید لطفا ابتدا وارد شوید، در غیر این صورت می توانید ثبت نام کنید.